



Short peptides and D-7 have stimulated proliferation-associated protein Ki-67 decreased during aging in tissue-specific cell cultures received from young and old animals and in dissociated vascular endothelial cell cultures. Peptides and D-7 have interacted with promoter region of MKI67 gene coding protein Ki-67 that was obtained using methods of molecular docking. Both peptides have contacted with core promoter b’-agcctcaaccatcaggaaaacaagagt-3 located in MKI67 (ENSG00000148773) gene from -14 to +12 base pairs relative to the transcriptional initiation site through sequence CATC. Thus, vasoprotective effect of peptide revealed previously in elder people could be realized through epigenetic regulation of Ki-67 gene expression.
Shot peptide which has a high biological activity against vascular and immune tissue. Peptide in combination with conventional treatment in patients of older, elderly and senile age with atherosclerosis of the arteries was more effective than using only standard therapy. Shot peptide administrated per os has contributed to the normalization of sleep, decrease in the manifestations of cardiac arrhythmias and ischemia of angina attacks in patients with coronary artery disease. In elderly patients with hypertension, oral administration of peptide in combination with antihypertensive therapy contributed to the achievement of long-term remission between hypertensive crises and the reduction of total cholesterol and Very low density lipoproteins in the blood [2]. The combination of oral administration of peptide and neuroprotective peptide pinealoma working at the age of 20-75 years of age in hazardous industrial conditions contributed to the improvement of the functions of memory, attention, thinking, accelerate perceptual-motor responses, improve mental health, decrease in the degree of aging of the CNS [1].
Well known that at the molecular-cellular level of age-related involution of the vascular system is associated with a reduction of intensity of processes of cell renewal of the endothelium [17]. It has been proved that with the aging of the vascular endothelium, its proliferative ability decreases, and the incidence of polyploid cells increases. In the human aorta, after 40 years old, the content of polyploid cells reaches 30 % of the total number of endothelial cells. This leads to a local disorder of the integrity of the endothelial layer and adhesion of monocytes with subsequent development of atherosclerotic vascular lesions [3]. There are some experimental data indicating that the vasoprotective properties of peptide are associated with its ability to regulate the expression of different signal molecules, including the transcriptional cell proliferation factor Ki-67.
Peptide stimulated the proliferation of human and animal immune and stem cells [4, 6, 12, 15]. In the study of the effect of peptide on T-helpers with CD4+CD25T’+ phenotype, it was found that it stimulated the expression of Ki-65 cell proliferation marker [13]. In the accelerated aging model, peptide restored the structure and increased the proliferative ability of rat duodenal cells by restoring the functional activity of its vascular component [3]. Recently, a peptide D-7 which according to conformational characteristics may have biological activity similar to shot peptide.
It is well known that the Ki-67 protein is a leading marker for evaluation of reduction of cell proliferative activity and degree of involutive processes in equal organs and tissues. Probably peptide stimulation of cell proliferation may be associated with activation of gene expression of proliferative protein Ki-67. One of the main factors of Ki-67 gene transcription regulating its expression is SP-1 protein. The last one interacts with the cow promoter of the Ki — 67 gene on two binding sites localized in the ranges from -159…-145 nucleotide pairs and from -14…+12 pairs of nucleotides (p. n.) with respect to the point of initiation of transcription [18]. These sites are responsible for the positive regulation of Ki-67 gene expression and are important for maintaining gene transcriptional activity. Earlier it was suggested that short peptides penetrate into the cell nucleus and site-specifically interact with the promoter sites of genes, epigenetically regulating their expression [5,10,14]. In addition, the possibility of interaction of tetrapeptide with DNA was shown in a foreign study by molecular modeling [7]. In this regard, the ability of peptide and D-7 to activate the synthesis of proliferative protein Ki-67 may be associated with their participation in the regulation of Ki-67 gene expression (MKI67).
The aim of this work is comparative study of the of peptides’ effect and D-7 on the expression of the proliferation marker Ki-67 in cultures of vascular endothelial cells upon aging and to assess the interaction of peptides with the promoter areas of the gene Ki-67 protein.
Materials and methods
Two types of cell cultures such as organotypic and dissociated have been used during the study. Organotypic cultivation of vascular tissue of young (of 3 months old) and old (24 months old) Wistar rats was carried out in Petri dishes in 3 ml of culture medium according to the previously described technique [11]. The nutrient medium included 35 % Needle medium, 35 % Hanks solution, 25 % fetal veal serum, 0.6% glucose, 0.5 u/ml insulin and 100 u/ml gentamicin. Vascular explants within 3 days were cultured in an incubator at a temperature of 37 °C under conditions of constant intake of 5 % CO2. Organotypic cultures of vascular cells were divided into three groups; control — with the addition of a physiological solution and two experimental — with the addition of peptide and D-7 at a dose of 0.05 ng/ml. cell Proliferation in organotypic vascular culture was investigated by lifetime light microscopy. The area index calculated as the ratio of the total area of the implant to the initial area (the area of the central zone) was determined to quantify the growth of the explants. The explants were photographed using a video camera for a microscope MTN-13 «Alfa-Telecom», series 10. Photom1.2 program was used to calculate the index of the Explant area.
By enzymatic dissociation using collagenase fragments of aorta of young rats Wistar line (3 months old) were obtained primary dissociated culture of endothelial cells. The nutrient medium for dissociated vascular cell cultures contained 15 % fetal bovine serum, 82.5 % DMEM, 1.5 % HEPES, L-glutamine and gentamicin. Times passage produced after 3 days on the 4th, when the culture reached 80% of confluence. Cultivation was carried out up to the 3rd passage, which was regarded as «young» cell cultures, and up to the 14th passage, which was the «old» endothelial cell cultures in accordance with the recommendation of the International Association for cell culture research (USA, San Francisco, 2007). To study the dependence of the biological activity of peptides on the concentration, dissociated cultures of endothelial cells were divided into three groups: control — with the addition of saline and two experimental — with the addition of shot peptide and D-7 at a dose of 20 ng/ml.
For immune cytochemical staining of cell cultures vessels were used primary monoclonal antibodies against Ki-67 (Novocastra, 1:50) and secondary antibodies biotinylated — wide anti-mouse immunoglobulin (Novocastra). Permeabilization was performed using 0.1 % Triton X100. The reaction was visualized using radish peroxidase and diaminoben — zidine (EnVision Detection System, Peroxidase/ DAB, Rabbit). Mouse). To evaluate the results of the reaction, a morphometric study was carried out using a computer analysis system of microscopic images consisting of a microscope «N)’:on Eclipse E400», a digital camera «Nikon DXM1200» and Videotest Morphology 5.2 software. The relative expression area was calculated as the ratio of the area occupied by immunopositive cells to the total area of cells in the field of view and expressed as a percentage
The MKI67 gene was analyzed to construct a computer model of peptide-DNA interaction (protein gene Ki-67) from the ENSEMBLE database numbered ENSG00000148773 and its promoter regions in the ranges from -159 to -145 BP and -14 to +12 BP with respect to the point of transcription initiation. The found promotor sequences were used to construct DNA sites and further study the interaction of peptides with these sites by molecular modeling methods.
Molecular modeling of DNA complex with peptides was performed using Molecular Operating Environment 2012 software [16]. To calculate the optimal mutual orientation of peptide and DNA molecules during their binding and formation of a stable complex, the Molecular Dokin method was used. The contact area, the number of hydrogen bonds, the parameters of hydrophobic and electrostatic interactions were taken into account in the calculations. We used Amber force field 12ЕНТ and genetic algorithm search GBV1/WSA. After the construction the DNA molecule and peptides was determined at pH 7 and T =300 K. The interaction energy of peptides with DNA was determined by the value of the estimated function 5 (kcal/mol), which was calculated by the formula:
S=c+a[2/3(ΔУ(coul)+ΔУ(sol))+ΔE(vdw)+βΔSA (weighted)
where C — the value of the loss of rotational and translational entropy of the complex; a, |3 — experimentally determined constants that depend on the force field; L ;— the value of the Coulomb energy, which is calculated using the system charge at a dielectric constant equal to 1; E , — the value of the electrostatic energy of the solvent; Evdw — van der waalson contribution to the interaction energy;-the contribution of molecular shells in the value of energy. 10 iterations of docking of peptide and D-1 with each promoter site were carried out. Solution docking the RA was rank in descending order from the most energetically favorable solution to the least energetically favorable. Of each docking solution (n=10), only the first solutions were analyzed, since they are the most energy efficient.
Statistical processing of experimental data included the calculation of the arithmetic mean, standard deviation from the mean and confidence interval for each sample and was carried out in the program Statistica 6.0. The Shapiro—Wilk criterion was used to analyze the type of distribution and test the null hypothesis. Non parametric procedures of single—factor variance analysis (kruskall-Wallis test) were used to estimate the statistical homogeneity of several samples. Differences between the groups were considered statistically significant at p<0.05.
Results and discussion
In organotypic cultures of vascular cells of young rats under the action of peptide, the area index significantly increased by 19 %, under the action of D-1 — by 22% compared with the control group. In organotypic cultures of vascular tissue of old rats the area index with addition of peptide and D-1 increased by 20%. Thus, peptides stimulated the growth of organotypic cultures of vascular cells obtained from young and old animals.
In organotypic vascular cultures obtained from old animals, the area of Ki-67 proliferation factor expression decreased by 72.5 % compared to cell cultures obtained from young animals. When injected into organotypic cultures of vascular cells of young rats, peptide increased the expression of Ki-67 proliferation marker by 12.5 % compared to the control group. When adding D-1 in culture cells of young animals, the area of expression of Ki-67 was increased by 15% (table 1). In organotypic cultures vessels of old rats under the influence of peptide and D-1 area of expression of Ki-67 increased, respectively, 1, and 2.2 times in comparison with the control group (see table. 1).
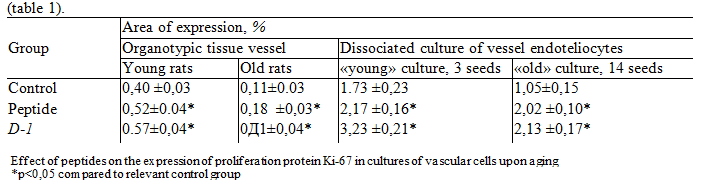
In dissociated cultures of endothelial cells, a decrease in the expression of Ki-67 marker in 1.64 times was observed during aging. The addition of peptide and D-7 to young cultures showed a significant increase in Ki-67 marker expression in 1.25 and 1.86 times, respectively, compared to the control group. When the peptide has been added and D-1 in the old cell culture expression of the marker Ki-67 increased, respectively, 1.97 and 2.02 times, compared with the control group.
To show the possibility of epigenetic regulation of gene expression of protein Ki-67 was created molecular models of the interaction of peptide and D-1 with the promoter portions of this gene in the ENSEMBL database were searched the promoter sequences of the gene Ki-67 protein. The MKI67 gene is localized on the 10th chromosome. Promoter sequences of three transcripts of MKI67 gene localized in different positions of chromosome were found. Each of the promoter ranges under study had three different sequences. So, in the range of -159…-145 BP. sequences 5 agcagttggcaagct-3′, 5’-agtcttcaaagaca-3 were studied relative to the transcription initiation point
and 5 -ccccgcccacgccct-W’. In the range of -14…+12 BP studied sequences 5’-acgctgcgggcg ggcgggcgggcggga-3’, 5-agcctcaaccatcag gaaaacaagagt-3’, 5’-ccaagagcacdauggaaaaggcccaa-3’. With the obtained sequences was performed docking of peptide and D-7. The results of docking are presented in table. 2.
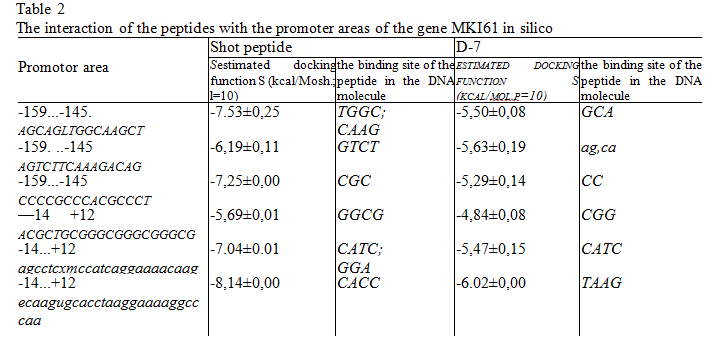
Analyzed the most energetically favorable solution to the docking of the peptides with DNA. Peptide and D-7 interacted with the DNA molecule, mainly from the small groove side. Only in one case, peptide contacted a large groove: in the promoter 5 ’-acgctgcgggcgggcgggcggg-3’ on the GGCG site, but such interaction was not among the most energy-efficient, and the interaction energy was -5.72 kcal/mol. When interacting with DNA peptide formed donor-acceptor and ionic bonds with 3-4 BP With all six promoter sites of the peptide were contacted on different sites, but with the lowest energy, he interacted with the promoter 5-ccaagagcacc taaggaaaaggccccia-3’ according to the website of SASS (see table. 2). The energy of formation of such a complex was 8.14 kcal/mol (Fig. 1, b). Due to the side group of lysine the vesugen acted as a donor of protons and formed a network of hydrogen bonds with oxygen atoms 04 02 deoxyribose and cytosine. Side chain carboxyl groups aspartic acid and C-terminal site, on the contrary, acted as acceptors of protons and formed hydrogen bonds with the amino group NH2 in the second position of guanine (see Fig. 1, b).
The peptide D-1 interacted in the DNA molecule with 2-3 BP With all promoters D 7 was contacted at different sites. However, as in the case of peptide, the most energy-efficient complex he formed with the promoter 5’ -ccaagagcacctaaggaaaaggcccaa-3’, but on another site — TAAS, located immediately after the binding site for peptide (see Fig. 1, a). The peptide formed hydrogen bonds with atoms 0’4 desoxyribose and 02 of thymine, however, the energy of interaction of peptide with DNA was higher and amounted to -6,02 kcal/mol. This difference in binding energies between peptide and D-7 is most likely associated with an additional functional group in peptide and, accordingly, with an increase in the area of contact with DNA in a small groove.
In experiments with organotypic tissue cultures vessels and dissociated cell cultures of endothelial cells established that both peptides increased the expression of the proliferation factor Ki-67, which was sniggles in aging cells of blood vessels (see table. 1). This allowed us to assume that the peptide and D1 may be a single binding site in the gene for the protein Ki-67. Such a site was found in the promoter 5’-agcctcaaccatcaggaaaacaagagt-3 (Fig. 2, a).
Both the peptide is contacted in the minor groove j site With a PBX and formed a hydrogen bond with the 02 of thymine (see Fig. 2, b, C). The binding energy differ by 1.5 kcal/mol, however, the D-7 was formed with DNA more hydrogen bonds than the peptide (see Fig. 2, b, C). Peptide formed one hydrogen bond with DNA between the amino group of the lysine side chain and the oxygen atom thymine. In the interaction of D-7 with a DNA molecule, a network of four hydrogen bonds formed between the amino group of the peptide backbone and the nitrogen atoms N3 of adenine and oxygen O2 of thymine and between the carboxyl group of aspartic acid and CH2 of the sugar phosphate backbone was found. In addition to hydrogen bonds, peptides formed hydrophobic, van der Waals and electrostatic interactions with DNA nucleotides, which were taken into account when calculating the estimated energy functions of the interaction of peptides with DNA. Therefore, to judge the profitability of the complex only by the number of hydrogen bonds formed is not entirely true.
Thus, the supposed molecular target of the action of peptide, whose vasoprotective effect was previously established in patients of older age groups with different vascular pathology, is the promoter zone of the gene MKI67 5’-agcctcaaccatcaggaaaacaagt-3’, encoding the proliferative protein Ki-67. This hypothesis is based on the increase of Ki-67 protein expression under the action of peptide in organotypic and dissociated vascular cell cultures during their aging. In addition, it was found that the D-7 had in the cultures of endotheliocytes upon aging effect similar to peptide. Earlier experiments with other short peptides showed the possibility of epigenetic regulation of expression of different genes [5, 8].
Similarly, the mechanism of action of peptide and D-7 was their binding to the promoter sites of the MKI67 gene (proliferative protein Ki-67 gene) responsible for the positive activation of its transcription. We were proposed the model of complexations of peptides with different binding sites of the gene МКI67 and calculated values of the energies of formation of these complexes.
It is established that peptide formed more energy-efficient complexes with the DNA molecule than peptide D-7. This is most likely due to the presence of an additional functional group in peptide. The promoter of the gene МКI67 5’-agcctcaaccatcaggaaaacaagagt-3′ shot peptide and D-7 was contacted with the DNA in one binding site of SATS Probably the interaction of both peptides with this website has identified the same effect of increasing the expression of proliferative protein Ki-67 peptides.
Conclusion
Thus, vasoprotective effect of peptide observed in people in older age groups with vascular pathology, probably associated with its ability epigeneticist regulate gene expression MKI67, which is a protein Ki-67 — activator of cell proliferation. In addition, another short peptide, D-7, according to molecular modeling and research in cell cultures, having an effect similar to peptide, can also be considered as a potential vasoprotective agent in age-related pathology of the cardiovascular system.
Literature
- Bashkireva A. S., Artamonova V. G., Peptidergic correction of neurotic conditions in drivers of trucks // Advance gerontology 2012. Vol. 25. No. 4. P. 718-728.
- Kitachev K. V., Sazonov A. B., Kozlov K. L. and others the Role of vasoactive peptide in the treatment of chronic arterial insufficiency of the lower extremities. Advance gerontology 2013. Vol. 26. No. 2. P. 292-296.
- Fingers M. A., Kvetnoy I. M., Polyakova V. O. and other Signal molecules: place and role in personalized diagnosis, treatment and prevention of socially significant diseases // Molecular honey. 2012. No. 5. P. 3-8.
- Anisimov V. N Khavinson V. Kh. Peptide bioregulation of aging: results and prospects // Biogerontology. 2010 Vol. 11. № 2. P. 139-149.
- Fedoreyeva L. /., Kireev 1.1., Khavinson V. Kh., Vanyushin B.F. Penetration of short fluorescence labelled peptides into the nucleus in HeLa cells and in vitro specific interaction of the peptides with deoxyribooligonucleotides and DNA // Biochemistry. 2011 Vol. 76. №11 P. 1505-1516.
- Grigor’ev E I., Khavinson V. Kh., Malinin V. V. et at. Role of aqueous medium in mechanisms underlying the influence of immuno active peptides in ultralow doses ,t Bui. exp. Biol. Med. 2003. Vol. 136. №2. P. 150-154.
- Huang H., Kozekovt.D., KozekovaA. etai. Minor groove ori- entationoftheKWKK peptidstetheredviatheN-terminal amine to the acrolein-derived 1 ,N2-gamma-hydroxypropanodeoxyguanosine lesion with a trimethylene linkage // Biochemistry. 2010. Vol. 27. № 49. P. 6155-6164.
- Khavinson V. Kh., Malinin V. V. Gerontological aspects of genome peptide regulation. Basel (Switzerland): Karger AG, 2005
- Khavinson V. Kh., Grigoriev E. I., Malinin V. V., Ryzhak G.A. Peptide, pharmaceutical composition, and a method of treating microcirculation disorders: US Patent 7. 851 449. 2010.
- Khavinson V.Kh., Solov’ev A. Yu., Tamovskaya S.I., Linkova N. S. Mechanism of biological activity of short peptides: cell penetration and epigenetic regulation of gene expression // Biol. Bull. Rev. 2013. Vol. 3. № 6. P. 451-455.
- Khavinson V.Kh., Linkova N.S., Pronyaeva V.E. et at. A method of creating a cell monolayer based on organotypic culture for screening of physiologically active substances II Bui. exp. Biol. Med. 2012. Vol. 153. № 5. P. 795-799.
- Khavinson V.Kh., Nikolsky I.S., Nikolskaya V. V. et al. Effect of tripeptides on lymphoid and stem cells // Bui. exp. Biol. Med. 2011. Vol. 151. № 6. P. 722-725.
- Khavinson V. Kh., Polyakova V. O., Linkova N. S. et at. Peptides regulate cortical thymocytes differentiation, proliferation, and apoptosis//J. Amino Acids. 2011. Vol. 2011. P. 1-5.
- Khavinson V.Kh., Tamovskaya S.I., LinkovaN.S. et al. Short cell-penetrating peptides: a model of interactions with gene promoter site // Bui. exp. Biol. Med. 2013. Vol. 154. Iss. 3. P. 403408.
- Lin’kova N. S., Polyakova V. O,, Trofimov A. V. et al. Peptidergic regulation of thymocyte differentiation, proliferation, and apoptosis during aging of the thymus // Bui. exp. Biol. Med. 2011. Vol. 151. №2. F 239-242.
- Molecular Operating Environment; Chemical Computing Group Inc (2012) 1010 Sherbooke St. West, Suite #910, Montreal, QC, Canada, H3A2R7.
- Olivieri F., Recchioni R., Marcheselli F. et al. Cellular senescence in cardiovascular diseases: potential age-related mechanisms and implications for treatment // Curr. Pharm. Des. 2013. Vol. 19. №9. P. 1710-1719.
- Pei D.-S., Qian G.-W., Van H. et al. Analysis of human Ki-67 gene promoter and identification of the Sp1 binding sites for Ki-67 transcription //Tumour Biol. 2012. Vol. 33. № 1 P. 257-266. doi:10.1007/s 13277-011 -0277-z. Adv. geront. 2014. Vol.27. №1. P.108-114